3.5. Система виділення
Видільна система комах складна, різноманітна і може бути підрозділена на три окремі групи органів або залоз – екстреторну систему, секреторну систему і ендокринну систему (рис. 11). Являється здатність виділяти на поверхню або в середину різноманітні речовини і тим самим брати участь в обміні речовин в організмі. В процесі переварювання їжі і використання речовин в організмі утворюють не потрібні або шкідливі для нього речовин. Ці не потрібні або шкідливі речовини, виділені з організму, називаються екскретами, а сам процес їх виділення – екскрецією. Головним органом виділення служать мальпігієві судини: названі вони так за ім’ям італійського вченого Мальнігі, який вперше відкрив їх у комах ще в ХVІІ ст. Це сліпі на вільному кінці трубочки, прикріплені своєю основою до кишечника на межі між середньою кишкою і задньою. Число мальпігієвих судин у різних комах коливається в межах 2-200 і більш або менш характерно для різних систематичних груп. Вільна частина мальпігієвих судин зважена і як би плавають в гемолімфі, відсмоктуючи із неї продукти виділення. Однак у гусениць метеликів, у багатьох жуків і їх личинок і у деяких інших комах кінець трубочок фіксував на задній кишці; це явище називається криптонефрія і забезпечує всмоктування трубочками води з задньої кишки, тим самим доповнює функцію останньої. Омиваючи мальнпгієві судини, гемолімфа віддає їм накопичені в ній продукти обміну. Ці речовини диффундирують в мальпігієві судини, з відкіля виводяться в шлунок і виділяються разом з екскрементами через анальний отвір. Екскреторна функція має також нижньогубні або лабіальні залози. Екскреторну накопичуючу функцію приписують також нефроцитам – групам клітин, здатним поглинати з порожнин тіла введені туди сторонні речовини: аміачний кармін, білки, хлорофіл та інші речовини. Видільним процесом вважають також і линьку, при якій організм звільнюється від азотистих речовин; в цьому випадку хітин і головні забарвлені речовини – мелоніни — повинні роздивлятися як екскрети. Різноманітні залози комах виділяють речовини, так чи інакше використані організмом. Такі речовини позначають секретами, а сам процес їх виділення називають секрецією. Розрізняють два основних типи секреції: ділення за допомогою типових залоз, і виділення безпосередньо в кров особливими залозами, другий тип залоз отримав назву ендокринних, а виділені ними речовини – гормони. Екдокринні залози різноманітні по функціях і будовою. Головну роль відіграють хімічні стимулятори, виділені комахами – феромони, або екзогормони. Виділені речовини називають гормонами, або інкретами, а сама секреція позначається як внутрішня секреція, або інкреція. Потрапляючи в кров, гормони транспортуються нею у всі частини тіла. В цілому ендокринні залози регулюють розвиток комах і пов’язані з ними явища – личинковий ріст, линьки, розвиток (діапауза), статеве дозрівання. Нейросекреторні клітини головного мозку виділяє мозковий гормон, функція якого різноманітна. В ряді випадків цей гормон контролює і активізує діяльність передньогрудних залоз; при припиненні його виділення у личинок і лялечок затримується ріст і розвиток; виникає діапауза. Передньогрудні , або проторакальні залози уявляють собою пару залоз, розташованих в черевній частині передньогруді з боків передньогрудного ганглія і зв’язаних з останніми нервами. Ці залози своєрідні личинкам багатьох комах і виділяють линочний гормон або екдизон. Екдизон припиняє діапаузу, визиває у личинки линьку і регулює розвиток личинки. Прилеглі, або додаткові тіла (corpora allata) розташовані над передньою кишкою позаду головного мозку, у вигляді пари округлих утворень. Вони виділяють ювенільний гормон або неотенин. Діяльність ювенільного гормону заключається у тому, що він сприяє розвитку личинкових органів і перешкоджає перетворенню в старшу фазу. Кардинальні тіла (corpora cordiaca) схожі за формою і положенню з прилеглими тілами, близько примикають до них, але розташовані спереду останніх, пов’язані нервами з головним мозком і личинкам, і імаго. Їх роль найменш вивчена. Ендокринні залози пов’язані один з одним, діють взаємопов’язано, але функціонують також під дією нервової системи і навколишнього середовища (тепло, світло та інші фактори). В цілому ці залози утворюють ендокринну систему, яка відіграє важливу роль в регуляції росту і розвитку комах (Бей – Биенко, 1966).
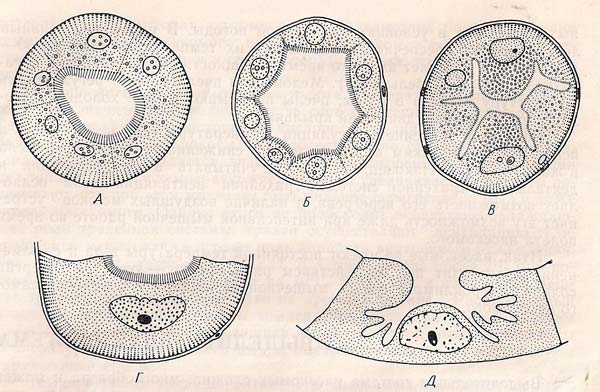
Перетинчастий розріз через різні мальпігієві судини:А-медведки; Б- Медоносної поли; В-плавунца,дистальна частина; Г і Д-лічинки звичайного мурашиного лева, різні клітини(Г — з основи Д — з верхньої частини)
3.6. Нервова система
Нервова система регулює усі функції організму, з’єднує його в єдине ціле і являється посередником між органами чуттів. Через органи чуттів організм сприймає інформацію з зовнішнього середовища, переробляє її в нервових системах і в співвідношенні зі своїм внутрішнім фізіологічним змістом здійснює необхідні цілеспрямовані дії. У комах нервова система сильно диференційована і підрозділена на центральну, периферичну і симпатичну нервову систему.
Центральна нервова система комах має метамерну будову, як і нервова система інших членистоногих і кільчатих червів. Основу її складає серія парних нервових вузлів, або гангліїв, з’єднаних продольними тяжами, або коннективами. Уся система гангліїв у комах підрозділена на два відділи – головний і черевний. Головний відділ складається з крупного надглоточного вузла, розташованого над кишечником і менш розвинутого підглоточного вузла. Вони з’єднані між собою тяжами, які опоясують передній відділ кишечника і утворюють навкологлоточне кільце. Черевний відділ складається з ганглії, розташований під кишечником в сукупність в черевний нервовий ланцюжок. Головний мозок у вигляді надглоточного вузла – головний центр всієї нервової системи комах. Він складається з трьох злитих комах – протоцеребрума, дейтоцеребрума і тритоцеребрума. Підглоточний вузол інервує ротові органи і передній відділ кишечника і походить в результаті злиття трьох гангліїв щелепного відділу голови – гнатоцифалона. Черевний нервовий ланцюжок складається у більш примітивних комах, з трьох грудних гангліїв і восьми черевних; в черевному відділі недостає трьох гангліїв, і останні сегменти не мають своїх нервових центрів. Пояснюється це переміщенням кінцевих гангліїв у 8 сегмент черева і злиттям їх тут в один загальний ганглій. Але тенденція до переміщення гангліїв у перед і до з’єднання їх більш великі ганглії проявляється і іншими частинами черевного нервового ланцюга, в результаті чого відбувається інтеграція нервових вузлів і зменшення їх чисельності. Скорочення числа гангліїв в черевному нервовому ланцюзі досягається шляхом об’єднання як черевних так і грудних вузлів, і в кінцевому рахунку може призвести до того, що усі вузли нервового ланцюга стають об’єднаними в два – три і навіть в один великий вузол, концентрація нервової системи у вищих груп – яскраве проявлення принципу олігомеризації, централізує і покращує нервове управління організмом в цілому, сприяє загальному підвищенню морфо – фізіологічного рівня комах. Вона може розглядатися еволюції комах також і як проявлення іншої еволюційної закономірності – принципу ароморфозу (рис. 12).
Периферична нервова система утворена з нервів, які відходять від гангліїв центральної і симпатичної нервової систем. За допомогою нервів центральна і симпатична нервові системи становляться зв’язаними з різноманітними органами. До периферичної нервової системи слідує також віднести розташовані по тілу чутливі нейрони, частіше зі вільними нервовими закінченнями. Симпатична нервова система, або вісцеральна — регулює роботу внутрішніх органів і м’язової системи комах. Вона підрозділяється на три відділи: рото – шлунковий, черевний і хвостовий. Рото – шлунковий відділ має найбільш складну будову і знаходиться над переднім відділом кишечника. Важливими його частинами являються лобний ганглій, лежачий над стравоходом з переду головного мозку, і відходячий від нього назад зворотний нерв, останній проходить під головним мозком і закінчується позаду нього шлунковим ганглієм з доповнюючими утвореннями. Рото – шлунковий відділ обслуговує наличник, верхню губу, а також область передньої кишки, серце і аорту; у деяких комах він керує ковтальними рухами. З цим відділом анатомічно зв’язані ендокринні залози – прилеглі і кардіальні тіла. Черевний, або вентральний відділ показаний непарним нервом, який простягається у вигляді тонкого тяжа між коннективами черевного нервового ланцюга з парою бокових відростків у кожному сегменті. Непарний нерв регулює роботу крилових і інших м’язів, і збуток його визиває швидку втому. Хвостовий, або каудальний відділ пов’язаний з заднім вузлом черевної нервової системи ланцюга і іннервує відділ кишечнику і статеву систему. Збудження і затримання складає важливу основу всієї нервової діяльності організму. Проведення збудження з однієї частини тіла в іншу здійснюється за допомогою нервів двох типів: в одних нервів воно іде від нервової клітини до м’язів або до іншого органу, в інших випадках має зворотній напрямок і йде від сенсорних клітин збудження органів чуттів до нервового центру. Збудження має електрохімічну будову і проявляється у вигляді серії швидких змін потенціалу, протікаючих в нейронах і нервах; збудження розповсюджується хвилеподібно. Затримання являється зворотнім процесом, але як показали дослідження російського фізіолога – Н.Є. Введенського (1852 р. – 1922 р.), збудження і затримання за своєю природою єдині, і надмірне збудження призводить до затримання. Затримання здійснюється за допомогою центрів затримання, які можуть розташовуватися як у головному мозку, так і в інших частинах центральної нервової системи. В останній час у комах виявляється також можливість периферичного затримання через непарний нерв симпатичної нервової системи. Збудження центрів затримання підвищує поріг рефлекторної відповіді, довго тривале затримання можливе лише при відсутності посилання стану, і для виводу з подразнення затримання необхідно посилення подразнення фактору.
Органи чуттів являються посередником між зовнішньою середою і організмом, органами інформації про стан зовнішніх умов життя. За аналогією з органами чуттів вищих тварин і людини у комах розрізняють органи дотику, слуху, нюху, смаку і зору. Розрізняють у комах наступні відчуття з їх рецепторами: механічне відчуття , слух, хімічне відчуття , гігротермічне відчуття і зір. Механічне відчуття представлене механорецепторами, які сприймають різноманітні механічні дії і представлені всього лише однією клітиною. Сюди відносяться дотичні рецептори, а також чуттєві структури, сприймаючи струс, положення тіла, його рівновагу.
Обов’язкові, або тактильні рецептори розкидані по всьому тілу у вигляді простих незанурених сенсил з з’єднаним чуттєвим волоском. Зміна положення волоска при торканні твердого предмета або під дією руху води або повітря передається чуттєвою клітиною, де і виникає збудження, яке передається по її відросткам в нервовий центр. Другий вид механорецепторів складає дзвоноподібні сенсили, які відрізняються від попередніх відсутністю чуттєвого волоска і зануреного у шкіру. Їх рецепторна поверхня має вид кутикулярного ковпака або дзвона і знаходиться на поверхні кутикули. До числа механорецепторів слідує віднести і хордотональні органи, нерідко роздивляючи їх як органи слуху. Однак у комах важко провести границю між сприйняттям механічних струсів і слуховим чуттям, сприймаючи тиск повітря. Хордотональні органи уявляють собою серію особливих сенсил, натягнутих між двома частинками кутикул. Хордотональні органи розташовуються на різноманітних частинах тіла – на черевці, вусиках, ногах, крилах і т.д. Розділені вони симетрично метамерно і зустрічаються в великому числі – десятками пар. Слух розвинутий далеко не у всіх комах. У прямокрилих, деяких клопів і ряді лускокрилих слухові рецептори представлені тимпанальними органами. Особливо гарно розвинуті вони у тих комах, які володіють звуковими органами і можуть видавати звук у вигляді стрекотання або співання.; аналогічно тимпанальні органи уявляють собою накопичення сколопофорів, які пов’язані з потовщенням у вигляді барабанної перетинки частинками кутикули. Зовні вони гарно виражені у прямокрилих. У денних метеликів тимпональні органи знаходяться на надутій основі передніх крил, у совок – між груддю і черевцем. Функцію органу слуху у комарів виповнюють також Джонстонов орган, на церках у деяких тараканів і прямокрилих і на тілі гусениць метеликів розташовані слухові волоски, які здатні уловлювати звукові хвилі. Значення органів слуху у комах двояке. З однієї сторони, з їх допомогою сприймаються сигнали, що йдуть від осіб свого виду, що забезпечує зв’язок статей. З іншої сторони, органи слуху уловлюють і інші звуки, що йдуть із зовнішнього середовища; деякі комахи реагують на різкий звук, деякі паразитичні види, можливо розшукують жертву по видавництву нею звуку. Хімічні чуття служать для сприйняття хімізму середовища, запаху, смаку, і представлене хеморецепторами, в ряді випадків уловити різницю між нюховими і смаковими рецепторами важко, цілеспрямовано говорити про органи хімічного чуття в цілому. Однак фізіологічна різниця між цими рецепторами заключається в тому, що нюх сприймає газоподібне середовище з низькою концентрацією речовини, а смак – рідке середовище з високою його концентрацією. Нюх служить комахам різноманітно – для розшуку статі, розпізнання осіб свого виду, для знаходження їжі і міст відкладених яєць. Багато комах виділяють речовини – статеві аттрактанти. Смак грає більш специфічну роль, ніж нюх, так як служить лише для розпізнавання їжі. Комахи розрізняють чотири головні смаки – солодкий, гірки, кислий і солоний. Смакові рецептори розташовуються на ротових частинах, але можлива і інша їх локалізація. У метеликів вони знаходяться на лапках ніг і знаходять велику чутливість; при доторканні підошвенної сторони лапок до розчину цукру голодний метилик реагує розгортанню хоботка. Висока ступінь розвитку у комах хімічного почуття являється суттєвою стороною їх фізіології і служить науковій основі при вишукування і примінення деяких прийомів хімічної боротьби з шкідливими видами. Гігротермічне почуття має суттєве значення в житті ряду комах і в залежності від умов вологості і температури середовища регулюють поведінку особи; воно також контролює водний баланс і температурний режим тіла. Зір разом із хімічним почуттям можливо грає провідну роль в житті комах. Органи зору мають важку будову і представлені двоякого роду очами: складними і простими. Складні, або фасеточні очі в числі двох розташовані з боків голови, нерідко дуже сильно розвинуті і тоді можуть займати значну частину голови (рис. 13). Кожне фасеточне око складається з багатьох зорових одиниць – сенсил, які називаються омматидіями, число їх в складному оці може досягати багатьох сотень і навіть тисяч. Омматидій складається з трьох видів клітин, утворюючих оптичну, чутливу і пігментну частину. Зовні кожний омматидій утворює на поверхні ока округлу або шестигранну ячейку – фасетку, від чого складні очі і отримали свою назву. Оптична, або преломлююча частина оммотидія складається з прозорого хрусталика і лежачого під ним також прозорого хрустального конуса. Хрусталик, або роговиця являється в сутності прозорою кутикулою, має вид двояковипуклої лінзи. Хрустальний конус утворений чотирма подовженими прозорими клітинами і разом з хрусталиком складає єдину оптичну систему – ціліндричну лінзу; довжина її оптичної осі перевищує її діаметр. Чуттєва частина розташована під оптичною, утворює світлові промені сітчатку, або ретину і складається з серії (6 – 13) ретинальних клітин. Ці клітини витягнуті вздовж омматидія, розташовується секторіально і утворюють обкладку його центрального стержня – зірової палички, або рабдома. За допомогою складних очей комахи розрізнюють форму, рух, забарвлення і відстань до предмету, а також поляризоване світло. Прості очі розташовані між складними очами на лобі і темені або тільки на темені. Вони маленькі, в числі трьох і розташовані трикутником. В наслідок свого положення у верхній частині голови вони не рідко називаються такождорсальними вічками. Вічка розвинуті далеко не в усіх комах, відсутні у багатьох двукрилих і мкткликів. У безкрилих, або короткокрилих комах вони також відсутні або рудиментарні. Їх функція недостатньо з’ясована. Встановлено, що у ряді форм фокус вічка лежить за чуттєвою частиною, тому сприйняття зображення в даному випадку не може бути. Разом з цим існує анатомічний зв’язок вічкових нервів з нервами складних очей. Присутність вічок у гарно літаючих рухомих форм і відсутність у багатьох безкрилих комах вказує на регулюючу рух функцію. Вони стимулюють складні очі, посилюючи їх фотокінетичну реакцію. Від дорсольних вічок слідує відрізняти бокові, або латеральні вічка, личинки комах з повним перетворенням. Ці вічка, названі також стеммами, розташовані на бокових частинах голови на місці, де у дорослих особин лежать складні очі. Число їх різноманітне в межах одного і того ж виду. Одні види мають всього лише по одному вічку з кожної сторони, у других число їх досягає шести і більш пар. При переході комах дорослий стан бокові вічка атрофіруються і замінюються складними очами. Стемми різноманітні за частинами будови, але для них характерна присутність кришталика. У гусениць метеликів є також к конус і розвинутий всього один рабдом, що робить такий глазок сходним з омматидієм складного ока. ( Бей – Биенко, 1966).
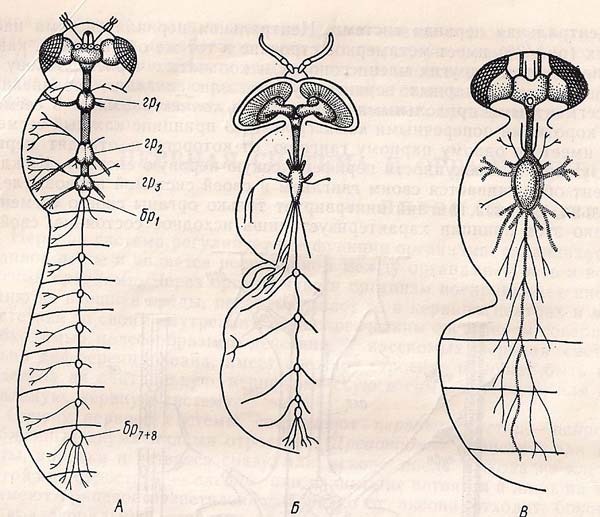
А – довговусі; Б – прямошові коротковусі; В – круглошові.
гр – грудні вузли; бр – брючні вузли.
3.7. Статева система.
Статева система виконує функцію розмноження і тим самим забезпечує життєдіяльність виду. Як правило, комахи роздільностатеві; самців прийнято позначати значком ♂, самок — ♀. Статевий диморфізм нерідко проявляється по ряду зовнішніх вторинно – статевих признаків – за формою і розмірами вусиків, величині тіла, різноманітних деталях будови. Статева система самця складається з пари гонад – семенників, пари семяпроводів, непарного семявиводного каналу, придаточних статевих залоз і чоловічого статевого придатку – едеагуса (рис. 14). Семянники мають вид округлих тіл і складаються з фолікулів, число та форма яких неоднакова у різних груп комах. В вершинній частині фолікула – гермарії – відбувається утворення живчиків, або сперматозоїдів; останні виникають шляхом багаторазових ділень з первинних семянних елементів – сперматогоній. Сперматозоїди, що утворилися з семянників поступють в семяпроводи, володіють особим розширенням – насіннєвими пузирями. Вони поступають в семявиводного канал, через нього досягають едегауса і при спарюванні з самкою виводяться на поверхню. Едеагус, або копулятивний орган – пеніс, утворює на поверхні геніталії самця.
Статева система самки складається з пари гонад – яєчників, пари яйцеводів, непарного яйцеводу, придаточних статевих залоз, семяприймача і нерідко яйцекладу. Яічники складають основу і найбільш сильно розвинуту частину статевої системи самки. Їх фолікули називаються яйцевими трубками, або оваріолами; число яйцевих трубок у різних комах – від 1-4 пар до 100 і більше, а у термітів навіть 2400 пар. За присутністю або відсутністю живильних клітин і розташуванню їх в яйцевих трубках (розрізнюють три типи яйцевих трубок). Паноістичний тип відрізняється відсутністю живильних камер, характеризує найбільш примітивний стан. Політропічний тип відрізняється присутністю живильних клітин, які чергуються з яйцевими клітинами, він характерний для рядів комах з повним перетворенням – сітчатокрилих, лускокрилих, перетинчатокрилих, двокрилих і м’ясоїдних твердокрилих. Телотрофічний тип, також – акротрофічний, характеризується розташуванням живильних клітин в вершинній частині трубки, звідки живильний матеріал поступає до місцевих клітин по протоплазматичним тяжам; він характерний прямокрилим. Дозрілі яйця з місцевих трубок поступають в парні яйцеводи, потім в непарний яйцевод і звідти через статевий отвір виходять наверх. В непарний яйцевод впадає тонкий протік семяприймачі; він має мішкоподібну форму і служить для зберігання сперматозоїдів, які попадають в нього при спарюванні. Запліднені яйця проходить при проходженні його через непарний яйцевод під час яйцекладки; сперматозоїди в цей час виходять з семяприймача і проникають в яйця. Придаткові залози відкривають свій проток також в непарний яйцевод і виконують різні функції – виділяють секрет для приклеювання яйця до субстрату, для обволакування груп яєць і утворення оптеки або кубишки. У статевого отвору нерідко розвинутий яйцеклад, який може бути і гарно видимим, або внутрішнім і скритим, або хибним. Дозрівання і відкладання яєць у кровосисних двокрилих не можливі без прийняття порції крові, тому до кровосисання яйця в яєчниках самки залишаються недорозвиненими.
Статева система комах має загальний план будови, але знаходить сильний статевий диморфізм. Особливістю будови являються важливою ознакою для розрізнення видів, родів і інших систематичних груп і широко використовується в систематиці комах (Бей – Биенко, 1966).

С – сімьяник; сп – сімьяпроводи; ж – придаточні статеві залози; ск – сімьяизвергательний канал; ко – копулятивний орган.
4. Особливості біології виду та родини, до якої належить вид капустяної совки
4.1. Тип метаморфозу і його сутність
Суть метаморфозу складається з того, стать що розвивається витримує на протязі життя суттєву перебудову своєї морфологічної організації і особливості біології. В зв’язку з цим з’являється диференціація постембріонального розвитку по крайній мірі на 2 фази – личинкову і дорослу, інакше названу імагінальна. У фазі личинки проходить ріст і розвиток особі, у фазі імаго – розмноження і розселення. В інших випадках між цими двома фазами з’являється проміжною фазою – лялечки.
Розрізняють два головних типи метаморфозу – неповне і повне перетворення.
Неповне перетворення характеризується проходженням лише трьох фаз – яйця, личинки і дорослої фази.
Капустяна совка моє повний тип перетворення ,при повному перетворенні весь цикл розвитку супроводжується проходженням 4 фаз – яйця, личинки, лялечки і імаго. Лялечки зовні несхожа з дорослою фазою, від чого вони й отримали назву; личинка завжди лишена фасеточних очей, видимих зачатків крил і часто мають інший тип ротових органів, ніж дорослі особи. Личинки комах з повним перетворенням живуть в інших умовах середовища, ніж дорослі. Більшість органів личинок цього типу мають тимчасовий, провізорний характер, виконують функції личинкового життя. До провізорних органів личинок комах з повним перетворенням відносяться: черевні ноги, ротовий апарат, шовковидільні, або павутинні залози (Бей – Биенко, 1966).
4.2. Форма яєць, розмір, колір і скульптура хоріона, способи відкладення яєць
Яйця совки шатроподібні, з приплюснутою основою, вершина їх має буро — червоним горбком, від якого по радіусам розходяться жолобки і реберця, колір відкладених яєць світло – жовтий, потім вони темніють; розташовуються яйця правильними рядами і 1 ярус, вся кучка має вид неправильного багатокутника (рис. 15). Діаметр яєць 0,6 – 0,7мм. Ембріональний розвиток яєць 4 – 12 днів.
Кладка яєць проходить кучками, на нижню поверхню листя капусти, салату (рис. 16). Кладка яєць починається з середини червня і продовжується до половини серпня, але бувають і сильні відхилення при несприятливій погоді. Кількість яєць в окремих кучках від 10 до 247, 30 – 80 яєць. Всього відкладають в середньому 1000 – 1500 яєць. При сприятливих умовах можуть відкладати максимально 2000 яєць (Литвинов, Євтушенко та ін., 2005).
4.3. Тип личинки, її характерні ознаки
Через 5 – 12 днів відроджуються личинка. Личинка капустяної совки типу справжньої гусениці, розміром 35 — 50мм, 16нога, мінливого забарвлення від сірувато – зеленого до темно – бурого, майже чорного, по бокам тіла широка жовта смуга, на спині – темний малюнок у вигляді «ялинки» (рис. 17). Найбільшої шкоди від гусениць першого покоління зазнають рання картопля та цибуля. Розвиток гусениць першого покоління триває 25 – 30 днів, після чого у ґрунті вони перетворюються на лялечок (на півдні – у 2 половині червня, у центральних і північних районах – у 1 половині липня). Гусениці 2 покоління пошкоджують середню і пізню капусту (рис. 18), а також столові буряки. Розвиваються вони повільніше, ніж гусениці 1 покоління, і здебільшого перетворюються на лялечок лише через 35 – 45 днів після відродження (Литвинов, Євтушенко та ін., 2005).
4.4. Тип лялечки і її характерні ознаки, в тому числі наявність захисних пристосувань
Лялечка капустяної совки червоно – бура, блискуча, нерухома, покрита, усі її придатки злиті з тілом (рис. 19). Розмір лялечки 19 – 24мм. У деяких нижчих груп лялечка вільна, з рухомими щелепами, за допомогою яких вона робить отвір у коконі і навіть вибирається на поверхню. Лялькування проходить не там, де жила і живилась гусениця, у багатьох видів – в ґрунті, відкрито розташований лялечці часто знаходяться в коконах, виготовлених з шовку з домішками інших речовин, рідше з чистого шовку (Васильєв, 1974).
4.5. Статевий диморфізм і поліморфізм у імаго
Як правило, комахи роздільностатеві; самців прийнято позначати значком ♂, самок — ♀. Статевий диморфізм нерідко проявляється по ряду зовнішніх вторинних – статевих ознак – за формою і розмірами вусиків, величині тіла, різноманітних деталях будови. Статева система самця складається з пари гонад – семенників, пари семяпроводів, непарного семявиводного каналу, додаткових статевих залоз і чоловічого статевого придатку – едеагуса. Семенники мають вид округлих тіл і складаються з фолікулів, число та форма яких неоднакова у різних груп комах. В вершинній частині фолікула – відбувається утворення живчиків, або сперматозоїдів; останні виникають шляхом багаторазових ділень з первинних семенних елементів – сперматогоній. Сперматозоїди, що утворилися з семенників поступють в семяпроводи, які мають особливі розширенням – насіннєвими бульбашками. Вони поступають в семявивідний канал, через нього досягають едеагуса і при спарюванні з самкою виводяться на поверхню. Едеагус, або копулятивний орган – пеніс, утворює на поверхні геніталії самця.
Статева система самки складається з пари гонад – яєчників, пари яйцеводів, непарного яйцеводу, допоміжних статевих залоз, семяприймча і нерідко яйцекладу. Яєчники складають основу і найбільш сильно розвинуту частину статевої системи самки. Їх фолікули називаються яйцевими трубками, або оваріолами; число яйцевих трубок у різних комах – від 1-4 пар до 100 і більше, а у термітів навіть 2400 пар. По закінченню розвитку яйця живлення його закінчується, фолікулярний епітелій виділяє хітиноподібні речовини і утворю зовнішну оболонку яйця – хоріон.
Найбільш розповсюджений тип поліморфізму – статевий поліморфізм. Для комах характерна диференціація статей в родині на декілька форм, самців, самок, робочих; останні являються нестатевозрілими самками, для позначення різних форм статей термін – каста. Кожна каста виникає в результаті статевих пристосувань і направленого до личинок (рис. 20)(Бей – Биенко,1966).
4.6. Життєвий цикл
. Весь цикл розвитку комахи, починаючи з фази яйця і закінчуючи дорослою фазою, досягнутої статевої зрілості, позначається поняттям покоління, або генерація, вона змінюється в значних межах і залежить від 2 головних факторів – спадкової ритміки життєвого циклу і вплив зовнішніх умов життя. Капустяна совка дає 2 покоління (табл. 1). Якщо після кладки яєць родина погибає, а сам період кладки був недовгий , то при декількох поколіннях з них чітко відмежовано від попередньої і наступної генерації. Але дуже часто тривалість життя імаго може бути значною, а період яйцекладки розтягнутим; в цих випадках покоління налягають один на одного і їх розмежування стає важким. Капустяна совка зимує на глибині 5 – 10см (Литвинов, Євтушенко та ін., 2005).
4.7. Діапауза
Діапауза уявляє собою стан фізіологічного спокою і виникає в життєвому циклі як спеціальне пристосування до переживання несприятливих умов в областях з сезонно – періодичним кліматом. В помірному кліматі виробляються пристосування до життя в умовах двох головних сезонів року – сприятливого літнього і не сприятливого зимового. При діапаузі проходить затримка росту і розвитку особі. Ця затримка виникає під дією ендокринної системи на основі нейро – гуморального регулюючого механізму; нервова система контролює діяльність ендокринної системи, а остання забезпечує взаємодію між органами через посредництво життя внутрішньої середи – гемолімфи. Личинки і дорослі особі, що знаходяться в діапаузі припиняють живлення, їх м’язова активність нерідко знижується і комахи становляться нерухомими або малорухомими. У личинок припиняється ріст і розвиток, в яйці припиняється розвиток зародку. Все це свідчить про різке зниження обміну речовин в діапаузному організмі. Показником служить зменшення швидкості дихання і падіння споживання кисню.
Настання діапаузи у активних фаз розвитку і лялечок завжди передує період інтенсивного живлення і накопичення запасних живильних речовин. В фазі яйця має місце ембріональна діапауза (Бей – Биенко, 1966).
5.Особливості екології виду
Екологія(від грецьких слів «ойкос» — дім, місце перебування та «логос» — вчення) – наука про відносини між рослинними та тваринними організмами також спільне такими з навколишнім середовищем.
Об’єктами екології можуть бути види, популяції організмів, спільноти та біосфера в цілому. Тому у наш час екологію поділяють на три розділи: екологію особи – аутекологію, екологію сукупностей осіб даного виду, що займають визначену ділянку території, — популяційну екологію та екологію біоценозів, тобто екологію сукупностей всіх видів організмів, що заселяють територію з більш або менш однорідними умовами існування, — біоценологію.
Вивчення впливу різноманітних факторів середовища на плодючість та витривалість(виживання) організмів, і як наслідок, їх вплив на зміну чисельності популяції у природі – є однією із найважливіших задач екологічних дослідів. Вивчення закономірностей коливання чисельності організмів дає можливість прогнозувати ці коливання та шукати шляхи змін умов існування у несприятливому для розмноження шкідливих організмів напрямку.
Екологічні фактори зазвичай розділяють на абіотичні, ґрунтові, біотичні та антропогенні. До абіотичних(або неорганічним)фактором відносять дію на організм кліматичних умов(температура, вологість, світло та інші), а також вплив магнітного поля землі, сили тяжіння, рівня радіоактивності та інше. За характером впливу на організм близький до абіотичних ґрунтові, або одафічні, фактори у зв’язку з різноманіттям частіше виділені в окрему групу. До біотичних, або органічних, факторів відносять особливості взаємовідносин між живими організмами переважно на основі харчування. Антропічні, або антропогенні, фактори складають особливу групу факторів, що виникли як якісно нове явище у житті нашої планети у зв’язку з інтенсивною господарчою діяльністю людини.
Що ж стосується екологічних особливостей виду – Ріпаковий пильщик, який я досліджую, можна вказати, що це фітофіл, тобто мешканець рослин(хорбіонт – мешканець густого травостою).
Можна зробити акцент на тому, що важливу роль у житті пильщика відіграють абіотичні фактори.
5.1. Абіотичні фактори
Абіотичні, або неорганічні фактори: дія на організм кліматичних умов(тепла, вологості, світла та інших умов), а також таких факторів, як сила тяжіння, склад атмосфери, радіоактивність, рельєф поверхні та інше..
5.1.1. Вплив температур на розвиток комах
Температура тіла комахи і його становище закономірно змінюється, при зміні зовнішніх умов. Активна життєдіяльність комахи можлива тільки в межах визначеного діапазону температур, який у різних видів різний. Вище чи нижче цієї температурної зони активності комахи втрачають здатність бути активними, впадаючи в стан теплового, або холодового заціпеніння, але при цьому зберігається здатність повернутись до активного стану, якщо температура середовища знову повернеться у межі зони активності. Дуже сильне підвищення, або зниження температури, що виходять за рамки фізіологічних можливостей особин, або виду викликає загибель організму (Щеголев,1960).
Швидкий розвиток комах у значній мірі залежить від температурних умов середовища. Розвиток комах може протікати тільки у рамках визначених температурних межах – верхній і нижній, що отримали назву пороги розвитку. Температури, що лежать вище нижнього порогу і ті, що не виходять за рамки верхнього порогу, отримали назву ефективних температур і тільки вони можуть забезпечити розвиток комахи, тобто тільки вони мають здатність викликати активізацію фізіологічних умов і надати ефект – виведення комахи із нульового становища. На основі даних суми ефективних температур та порогу розвитку можна приблизно визначити проміжок розвитку при тих чи інших постійних температурах (Бей – Биенко,1966).
Таблиця 2
5.1.2. Залежність розвитку комахи від вологості середовища
Показники вологи середовища різноманітні, але в екології комах найбільше значення має відносна вологість повітря, тобто відсоток насичення його водними парами. Дія вологи впливає різноманітно: вона має склад води у тканинах комахи, а у зв’язку з цим впливає на її поведінку, її виживання і плодючість. В одних випадках вологість середовища діє безпосередньо, в інших вона діє через їжу, яка вживається комахою, або через інші фактори середовища.
Маючи малі розміри і велике поверхневе випаровування, комахи стають залежні від вологи середовища, як екологічного фактору. Утримання в організмі вологи у потрібних межах вимагає спеціальних механізмів, що є особливими пристосуваннями для підтримки потрібної рівноваги між організмом і середовищем. Такими пристосувальними організмами, являються морфологічні, фізіологічні та екологічні адаптації. До числа морфологічних пристосувань відносяться: розвиток на покривах водонепроникної епікутикули, потовщення кутикули, будова дихалець, будова лялечки.
До числа фізіологічних механізмів регулювання водного балансу тіла комахи відносяться такі пристосування як відсмоктування води задньою кишкою із не перетравлених залишків їжі, поглинання вологи покривами при контакті з нею, отримання вологи в організм з їжею. Але за ступенем потреби до вологості середовища комахи неоднорідні, проявляють різноманітну вибірковість і серед них можна розрізнити три групи видів: надто вологолюбні, або гігрофілів, середньовологолюбні, або мезофілів і сухолюбні, або ксерофілів (Бей — Биенко,1966).
5.1.3. Спільна дія температури та вологості середовища на комах
Температура середовища у найбільшому ступені може змінювати характер дії вологи на комах. Тому необхідно досліджувати спільну дію температури та вологості середовища на швидкість розвитку, стан і плодовитість комах.
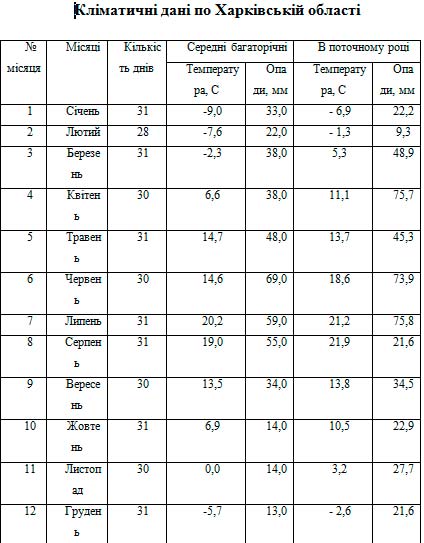
Всі основні показники життя і становища комахи – смертність, тривалість розвитку і плодовитість залежать у значній мірі від конкретного співвідношення температури і вологості середовища. Для оцінки і аналізу впливу температури і вологості середовища на комах у природній обстановці застосовується спосіб клімограм. Клімограма природніх умов Харківської області зображена на малюнку (рис, 21, табл. 3).
5.1.4. Роль світла у формуванні річного і сезонного циклів розвитку комахи
Роль світла як екологічного фактора у житті комах велика і часто не поступається, а у деяких випадках навіть переважає роль інших кліматичних факторів – температури і вологи. Світло виступає як фактор, що регулює річний життєвий цикл видів: довгий фотоперіод сприяє безперешкодному розвитку багатьох видів, тоді як короткий фотоперіод, який наступає у кінці літа – на початку осені, стимулюють перехід у стан діапаузи. Короткий фотоперіод точним і безпомилковим космічним сигналом скорого атакування несприятливого осіннього – зимового періоду життя і перехід у стан діапаузи, за цим сигналом забезпечує виду своєчасну фізіологічну перебудову для підготовки до зими.
Виявилось також, що фотоперіодична реакція комах не тільки при сильному, але і при слабкому освітленні: численні види чутливі до сили світла. Висока чутливість до світла має пристосувальне значення і завдяки їй виключається вплив на фотоперіодичну реакцію коливань інтенсивності денного світла при зміні погодних умов, наприклад у безхмарні сонячні і похмурі періоди дня.(Талинський,1988).
Дія світла відображається не менше у тому, що світло відіграє переважаючу роль у формуванні річного і сезонного циклі розвитку. Світло впливає на життя комах і по іншому. Передусім комахи розрізняються різною життєдіяльністю протягом світлової і темної частини діб: одні види активні при денному світлі, інші – при сутінках, треті – у ночі.
Суттєвою є також реакція на штучне світло нічних, сутінкових і денних комах. Як виявилось, багато комах, особливо активно, летять уночі на ультрафіолетове випромінювання.(Бей — Биенко,1966).
5.1.5. Значення повітряних потоків у розселенні виду
Суттєве значення у розселенні комах відіграє фактор повітряних потоків – горизонтальний і вертикальний. Велика кількість комах не може летіти, коли протистоїть вітру і тому мають пасивний політ(метелики, попелиці, дрібні двокрилі ті інші). Вітер, як горизонтальний потік повітряних мас є для таких комах, одним з найважливіших факторів розселення.
Переміщенню комах допомагають також і вертикальні повітряні потоки, наприклад: рух теплого повітря вверх, вдень, та вниз, увечері. Ці повітряні потоки підхвачують комах, переміщують їх у більш високі горизонти, де вони під дією сильних і постійних вітрів і повинні дрейфувати у повітрі.
5.2. Гідрофітні фактори
Гідрофітні, чи водногрунтові фактори: дія води в ґрунті як особливих середовищ життя організму.(Бондаренко, 1983)
Більшість крилатих комах в активних фазах свого розвитку, особливо у зрілій фазі, відносяться до числа тих, що живуть у повітряному середовищі. Значна частина цих крилатих у неактивних фазах свого розвитку(яйце і лялечка), а іноді, також, у фазі личинки є жителями прісних вод і ґрунту. Окрім цього, більшість первинно безкрилих комах і частина крилатих відносяться до числа постійних поселенців ґрунту у всіх фазах розвитку.
У цілому вище 90% видів усіх комах у той чи інший період свого життя або ґрунтом як середовищами життя і тому є частиною або повністю водяними або ґрунтовими мешканцями. Навіть у таких типових мешканців повітряного середовища, як сарани, метеликів, мух та інших активних літаючих комах, значну частину життєвого циклу проводять у ґрунті.
5.3.Біотичні фактори
Біотичні, або органічні фактори: дія на організм живих сил природи, взаємовідношення між організмами на основі живлення , внутрішньовидові відношення та інше.
Основу взаємовідношення комах з біотичними факторами середовища складають місцеві, або трофічні, взаємовідносини і зв’язки. Споживання їжі являється фізіологічною необхідністю, що і викликає появу різноманітних пристосувань, які направлені для використання тих чи інших джерел їжі.
Так як комахи – гетеротрофні організми, вони потребують органічні речовини, які утворені іншими живими істотами. Азот і вуглевод тіла комахи запозичують у процесі живлення від інших тварин і рослин, що утворює найтісніший зв’язок комахи-живителя від організмів-постачальників їжі (Бей — Биенко,1966).
5.3.1. Екологічний зв’язок комах із рослинами
Джерела їжі комах різноманітні. В одних випадках для живлення і отримання їжі використовуються живі організми – тварини і рослини; в інших випадках використовуються продукти їх життєдіяльності – неперетравлені залишки їжі у вигляді посліду і відмерлі частини рослин і можуть використовуватись в їжу самі тварини або рослини.
Спеціалізація видів комах на цих джерелах їжі сприяла виникненню різноманітних травних режимів. Так виникли рослиноїдні комахи, чи фітофаги, до яких відносять велику кількість видів.
Фітофагія розглядається як прояв травної спеціалізації першого порядку, яка основана на використанні в їжу різних джерел органічної речовини. Окрім фітофагів існують хижаки, паразити, сапрофіти, некрофаги.
Паразити використовують інших тварин, не лише, як джерело їжі, але й як середовище мешкання. До них відносять багато перетинчастокрилих, двокрилих. Некрофаги використовують у їжу трупи тварин.
Іншою формою являється травна спеціалізація другого порядку: вона характеризує ступінь потреби до їжі у межах кожної категорії першого порядку. Порівняно до рослиноїдних комах розрізняють однорядних, або монофагів, тобто ті, що живляться будь-яким одним видом рослин чи не багатьма близькими видами; обмеженоїдних, або оліфагів, тобто ті, що живляться (багаточисельними) систематично близькими видами рослин, що відносяться до різноманітних родин. Види, що живляться різноманітною їжею тваринного і рослинного походження, називають всеїдними.
Капустяна совка є геофілом (Бей — Биенко,1966).
5.3.2.Взаємодія з іншими організмами
Взаємодія комах з природними ворогами, а особливо: патогенними організмами, паразитичними черв’яками, хижими і паразитичними комахами та іншими членистоногими, хижими хребетними тваринами. Всі вони відіграють ту чи іншу роль в обмеженні розмноження і розповсюдженні шкідливих комах, тому приносять користь людині і деякі з них використовуються для біологічної боротьби із шкідниками, але ці живі організми при ураженні корисних комах можуть причиняти людині суттєву шкоду (Захваткин,1986).
5.4.Антропогенні фактори
Господарська діяльність людини впливає на організми, у тому числі і на світ комах. Ця діяльність у наш час одна із найшкідливіших у природі.
Освоєння цілинних земель під посіви сільськогосподарських культур, вирубка та викорчовування з цією ціллю місць, висушування боліт, зрошення засушених степів, пустель, пасовища худоби і багато інших проявів господарської діяльності людини у великій мірі змінюють фауну комах. Одні види при цьому повністю витісняються і зникають, інші навпаки отримують сприятливі умови для життя.
Серед агротехнічних заходів великий вплив на видовий склад комах і чисельність особин чинить використання хімічних і біохімічних заходів обробітку ґрунту, сівозміни та інше (Бей-Бієнко, 1966).
Висновок
Ентомологія – наука про світ комах.
Капустяна совка (Mamestra brassicae) – належить класу комахи, підкласу крилаті або вищі комахи, відділу комах з повним перетворенням, ряду лускокрилі, родини совки. Метелик з розмахом крил 40 – 50мм, має 2 пари крил , верхні більші ніж задні. Ноги ходильного типу, формула лапок 5 – 5 – 5. Голова гіпогнатичного типу, вусики булавоподібні, ротовий апарат сисного типу.
Система травлення має 3 відділи: передню, середню та задню кишки. Передня кишка складається з глотки, стравоходу, зобу, м’язового шлунка. Середня кишка не підрозділяється на відділи і називається шлунком. Вона виконує різні функції : виділення ферментів за участю живлення, всмоктування продуктів живлення та проштовхування неперетравлених залишків їжі у задню кишку. Задня підрозділяється на тонку, товсту та пряму кишку.
Кровоносна система не замкнута, кров заповнює порожнину тіла між органами, омиває їх і тільки частково замкнута в особливий орган кровообігу – спинний.
Дихальна система складається з великого числа поверхневих трубок – трахей, які пронизують все тіло.
Видільна система підрозділена на три окремі групи органів або залоз – екскреторну систему, секреторну систему та ендокринну систему.
Нервова система комахи регулює всі функції організму, з’єднує в єдине ціле і є посередником між органами чуттів.
Статева система призначена для розмноження і тим самим забезпечує життєдіяльність виду.
Капустяна совка має повний цикл розвитку : яйце – личинка – лялечка – імаго.
Яйце капустяної совки має шатроподібну форму, колі світло – жовтий, діаметр 0,6 – 0,7мм. Ембріональний розвиток 4 – 12 днів. Гусениця розміром 35 – 40мм, колір темно – бурий або майже темний. Лялечка капустяно червоно – бура, блискуча, не рухома, усі її придатки злиті з тілом.
Капустяна совка роздільностатева.
Капустяна совка в рік дає 2 покоління.
Капустяна совка зимує у стадії лялечки.
Екологія – вивчає взаємозв’язок організму з навколишнім середовищем і наслідки цих взаємозв’язків. Капустяна совка є геофілом.
До родини совки належить дуже багато видів серед них : озима совка, оклична совка, совка – гама, горохова совка та інші види, які є шкідниками сільськогосподарських культур.
З агротехнічних заходів боротьби з капустяною совкою велике значення має: глибока зяблева оранка полів з-під капусти, цукрових буряків, гороху та інших просапних, при якій знищується чимала кількість лялечок; весняні культивації, міжрядний обробіток капусти та інших просапних культур під час заляльковування гусениць капустяної совки першого покоління (червень – липень); знищення бур’янів, нектаром яких живляться лялечки, а листками – гусениці совки. При пошкодженні капусти та інших овочевих культур гусеницями капустяної совки рослини обприскують хлорофосом у 0,3% концентрації (1,2-1,8 кг/га), 0,3 – 0,4 процентною суспензією, 30% метафосу (1,5 – 2,0 кг/га), 0,2 – 0,3% трихлорметафосу 3 (1,0 – 1,5 кг/га). Обробку рослин фосфорорганічними препаратами можна проводити до і після зав’язування головок і припиняти за 20 днів до збирання врожаю, трихлорметафосом 3 — до зав’язування головок. Застосування отрут найбільш ефективне проти гусениць молодших віків. У період масового відродження гусениць добрі результати дає фосфамід у 0,2% концентрації (0,8 – 1,0 кг/га). Його можна застосовувати за 30 днів до збирання врожаю. Велике значення в системі заходів боротьби з капустяною совкою має біологічний метод з використанням яйцеїда трихограми. Випускати трихограму на капусту та інші хрестоцвіті треба в 2 – 3 прийоми на початку і під час масового відкладення яєць з розрахунку 20 – 60 тисяч яйцеїдів на гектар літа.
Біологічні методи боротьби – це випуск трихограм на початку літа метеликів і повторне через 7 – 8 днів по 20 -30 тис. особин 1га , при кількості більше 80 яєць на 1м квадратному норму випуску трихограм збільшують відповідно до 40 – 50 тис. на 1га. В літку метеликів другого покоління трихограму випускають повторною.
Список використаної літератури
- Бей – Биенко Г.Я. Общая энтомология . — М.: Высш. школа, 1966. — 500с.
- Бондаренко Н.В., Глущенко А.Ф. Практикум з загальної ентомології.-Л.: Колос, 1972. — 352с.
- Бондаренко Н.В., Поспєлов С.М., Персов М.П.Общая и сельскохозяйственная ентомологія. – М.: Колос, 1983. – 496с.
- Васильев В.П. и др. Вредители сельскохозяйственных культур и лесных насаждений. – К.: Урожай, Т. 2. – 1974. – 608с.
- Захваткин Ю.А. Курс общей энтомологии. – М.: Агропромиздат, 1986. — 320с.
- Литвинов Б.М., Євтушенко М.Д., Байдик Г.В. та ін. Сільсьгосподарська ентомологія. – К.: Вища освіта, 2005. – 511с.
- Мигулин А.А., Осмоловский Г.Е. и др. Сельскохозяйственная энтомология. – М.: Колос, 1976. – 452с.
- Талинский В.М. Биологические основы вредности насекомых. – М.: Агропромиздат, 1988. – 195с.
- Щеглов В.Н. Сельскохозяйствення энтомология. – М.: Сельхозиздат, 1966. – 511с.