
Сорта интенсивного типа обладают сложным комплексом хозяйственно ценных признаков и свойств, главными из которых являются большая потенциальная урожайность, хорошая отзывчивость на удобрения и другие факторы улучшения плодородия почвы, комплексная или групповая устойчивость против вредителей и болезней, пригодность для выращивания при интенсивных энерго- и ресурсосберегающих технологиях. Кроме того, сорта этого типа практически в такой же мере, как и полуинтенсивные, должны отвечать требованиям в отношении качества продукции, зимостойкости, засухоустойчивости и т. д. Устойчивость против полегания, которая, в первую очередь, зависит от высоты растения, является важнейшим свойством сортов интенсивного типа. Если средне- и высокорослые сорта по другим параметрам отвечают требованиям интенсивных сортов, то их можно успешно выращивать на высоком агротехническом фоне, на суходоле и в условиях орошения только с применением ретардантов. Большинство сортов высокоинтенсивного типа относится к короткостебельным формам. В европейских странах в связи с повышением уровня земледелия каждые 50 лет высота растений пшеницы как генетическое свойство уменьшалась примерно на 15 см. В таком направлении шли сортосмены.
Взгляды на преимущества короткостебельных сортов развивались в чрезвычайно сложных противоречиях и в острой полемике селекционеров и земледельцев-практиков. Первоначально идея использования короткостебельных форм пшеницы для выращивания при хороших агротехнических условиях, которые в последующем стали определяться как интенсивные технологии выращивания, имела немного сторонников. В последующем эта идея стала очень популярной.
В Индии, Мексике и других странах, особенно в регионах с обширными площадями орошаемых земель, обычные сорта пшеницы были заменены на сорта-олукарлики, созданные главным образом в Международном центре по улучшению кукурузы и пшеницы (СIММYТ) в Мексике под руководством известного селекционера Нормана Борлауга. После этого пшеница по валовым сборам зерна вышла на первое место в мире. В Индии, например, с 1969 до 1979 г. валовой сбор пшеницы удвоился. В нашей стране во многих районах озимопшеничной зоны сеют также сорта, которые несомненно должны быть отнесены к низкорослым — типа Безостой 1. Широкое использование этой пшеницы в качестве исходной формы для гибридизации повлияло на формирование такого же сортотипа во многих странах Европы и других континентах. Настоящие полукарликовые сорта у нас стали выращивать позже, чем в ряде других стран. Связано это прежде всего с неблагоприятными условиями — частые почвенные и воздушные засухи, воздействие экстремальных факторов в период зимовки озимых культур и др. Полукарликовые сорта зарубежной селекции в своем абсолютном большинстве нельзя выращивать в основных районах распространения озимой и яровой пшеницы из-за слабой зимостойкости и недостаточной устойчивости против почвенной засухи, особенно в начальные периоды онтогенеза.
Во Всесоюзном селекционно-генетическом институте, Краснодарском научно-исследовательском институте сельского хозяйства им. П. П. Лукьяненко, Донском селекцентре, Украинском научно-исследовательском институте растениеводства им. В. Я. Юрьева, НПО «Днепр», НПО «Селекция» и других селекционно-опытных учреждениях страны созданы отечественные полукарликовые сорта озимой пшеницы мягкой и твердой, которые успешно прошли государственное сортоиспытание и их выращивают в производстве. Генетический контроль признака высоты растения у пшеницы мягкой и твердой имеет сложную природу. Многие селекционеры, работающие с короткостебельными формами, очень часто оперируют понятиями одно-, двух и трехгенные карлики, зачастую используя их лишь для определения степени различия по высоте растения. В действительности же признак высоты растений контролируется значительно большим количеством генов, особенно это относится к так называемым малым генам. Генетическая природа признака высоты растения у пшеницы изучена еще недостаточно. Ряд генов карликовости, хорошо известных генетикам, например, гены S и С, носителями которых являются виды пшеницы Triticum sphaerococcum и Т. compactum, мало или совсем не используются в селекции, так как с их функцией связано ухудшение ряда хозяйственно ценных свойств. Не увенчалась успехом и многократная попытка использовать доминантные гены карликовости сорта Том Пус из Тибета. Неперспективная для селекции и так называемая «травянистая гибридная карликовость».
Освещая исторический и генетический аспекты селекции полукарликовых форм пшеницы, многие авторы обращаются к характеристике японского сорта Норин 10. Широкое использование во многих странах этого сорта в качестве донора генов карликовости при селекции короткостебельных сортов знаменует собой новый этап в селекции озимой и яровой пшеницы. Интересно отметить, что в происхождении Норин 10 принимала участие и наша Крымка, завезенная первоначально в США, а затем в виде сорта Туркеи, выведенного из нее индивидуальным отбором.
Американский генетик V. А. Johnson с соавторами установил, что признак карликовости у Норин 10 контролируется тремя главными генами. Однако полностью генетическая природа этого сорта еще не изучена. Мало известно и о типах взаимодействия генов, вызывающих укорочение стебля.
На основе использования генов карликовости этого сорта создано наибольшее количество короткостебельных сортов во многих странах мира. В Международном центре по улучшению кукурузы и пшеницы создано большое количество полукарликовых и низкорослых сортов яровой пшеницы, которые широко распространились в Мексике, Индии, Пакистане и других странах.
Сорта озимой пшеницы фото:





Не меньшее значение для селекции имеет ген карликовости, который вместе с короткостебельными генотипами завезен из Японии. Он более 60 лет успешно используется в селекции европейскими селекционерами. Особенно распространился он в сортах итальянской и югославской селекции. Этот ген, хотя и происходит от японских сортов, как и три гена сорта Норин 10, однако отличается от них по ряду свойств. В свое время он был привнесен от Кляин 33 в Безостую 4, а затем в Безостую 1.
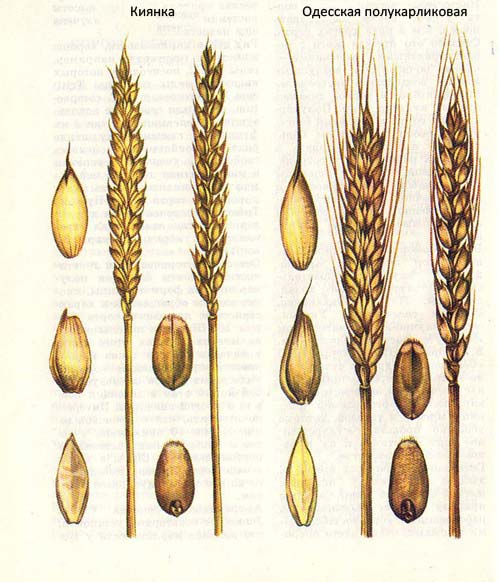
В нашей стране в селекции полукарликовых и низкорослых сортов озимой пшеницы широко применяются экспериментальные карликовые мутанты. При этом в одних случаях они послужили непосредственным исходным материалом для создания сортов (Киянка), в других — являются генетическими донорами короткостебельности для получения исходного материала методами гибридизации.
Генетическая природа признака короткостебельности у многих мутантов еще не изучена. Краснодарским научно-исследовательским институтом сельского хозяйства им П. П. Лукьяненко совместно с Институтом физической химии АН СССР получена из сорта Безостая 1 полукарликовая мутантная линия, которая под названием Карлик 1 была рекомендована академиком П. П. Лукьяненко в качестве источника для скрещивания при селекции короткостебельных сортов озимой пшеницы. Рекомендации П. П. Лукьяненко оказались весьма удачными. В Краснодарском НИИСХ на основе Карлика 1 созданы сорта озимой пшеницы Полукарликовая 49, Эстафета, Криница, Спартанка и др.
Во ВСГИ на основе этого же источника получены сорта Одесская 75, Лан, Прогресс; в Молдавском НПО «Селекция» — Питикул; в УНИИРСиГ им. В. Я. Юрьева — Полукарлик 3; в УНИИОЗ — Мрия Херсона. К. С. Бессараб и Е. Г. Жиров на основе моносомного анализа установили, что признак короткостебельности у Карлика 1 контролируется двумя генами. Выводы этих исследователей подтверждаются гибридологическим анализом. Правда, при этом анализе четко обнаруживает себя один эффективный ген рецессивного или слабо доминантного типа. Он возник в процессе мутации. Второй менее эффективный чисто рецессивный ген присутствовал в составе генотипа Безостой 1 еще до воздействия на нее мутагенных факторов. При скрещивании Карлика 1 с Безостой 1 менделирует только один ген, возникший в процессе экспериментального мутагенеза. Второй расщепления не дает, так как находится в гомозиготном состоянии.
Если не считать гены Д1, Д2, Д3, Д4, комплементарный эффект которых вызывает гибридную «травянистую карликовость», то можно сделать общее заключение, что все встречающиеся короткостебельные формы пшеницы содержат один, два или три гена карликовости (rht1; rht2; rht3). Однако общее количество генов, встречающихся в разных . генотипах, значительно большее. Их идентификация еще не закончена. Можно лишь отметить, что гены карликовости, которые происходят от Норин 10, отличаются от генов, содержащихся в ряде короткостебельных сортов европейской селекции (Сава, Русалка, Златна долина, Ардито, Бригантина и др.). Мутантный ген Карлика 1 отличается от тех и других генов, но зато второй ген этого источника такой же, как у некоторых европейских сортов. Гены карликовости сорта Том Пус совершенно отличаются от всех других, встречающихся в практически ценных сортах. Индийский селекционер, известный специалист по сортам полукарликового типа К. С. Гилл (1984), в особую группу выделяет гены карликовости сорта Олесон дварф и считает их непригодными для селекции сортов полукарликового типа. Исследования, проведенные во ВСГИ, показали другие результаты. Скрещивание Олесон дварф с озимыми сортами позволяет получать зимостойкие короткостебельные формы с отличным качеством зерна и хорошей устойчивостью против грибных болезней стебля и листьев. Кроме выше описанных генетических источников короткостебельности, в природе, по-видимому, существуют и другие еще не обнаруженные и не изученные носители генов карликовости. Так, среди ранее выращивавшихся на юге нашей страны был короткостебельный сорт яровой пшеницы твердой Хоранка. Признак короткостебельности от этого сорта легко передавался при гибридизации пшенице мягкой. Генетическая природа этого источника осталась неизученной.
Все основные гены карликовости локализованы в хромосомах геномов А, В, поэтому признак короткостебельности легко передается при межвидовой гибридизации от пшеницы мягкой к твердой, а также другим видам этого рода с таким же геномным составом.
Для селекции короткостебельных сортов пшеницы и разработки ряда агротехнических приемов их выращивания, в частности при определении оптимальных доз применения гербицидов, весьма важное значение имеет знание физиолого-биохимической природы признака карликовости.
Английский исследователь М. Галл с сотрудниками провели разнообразные исследования по выяснению физиолого-биохимической природы функции генов
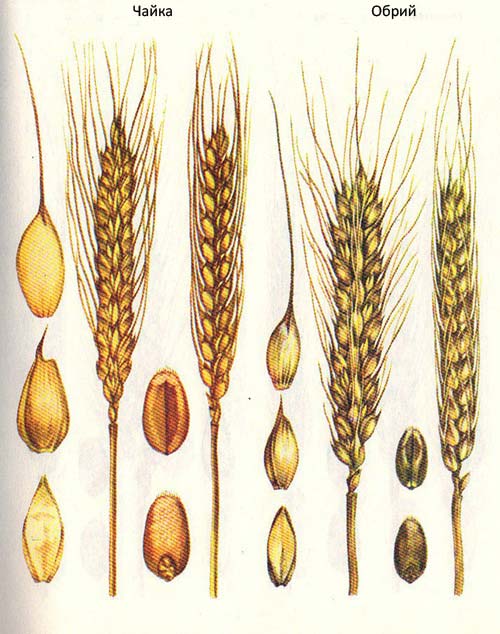
карликовости у ряда короткостебельных сортов и форм пшеницы мягкой. Установлено, что в одних случаях гены карликовости блокируют синтез ростовых веществ, поэтому растения в период стеблеобразования растут медленнее и в результате остаются низкими. Если такие генотипы (югославский сорт Сава) в период роста обрабатывать гиббереллином, то экзогенные вещества в значительной степени восполняют физиологический эффект, который осуществляется обычно веществами эндогенного происхождения, и растения будут расти быстрее. Другие гены карликовости не затрагивают процессов синтеза ростовых веществ в растении, а блокируют их утилизацию и таким образом исключают их из процессов метаболизма, отчего ростовые процессы сильно тормозятся. При скрещивании короткостебельных форм, относящихся к двум разным типам по реакции на ростовые вещества, наблюдается весьма интересное генетическое явление — гибридное потомство Е, значительно превосходит по высоте растения обоих родителей и не относится к короткостебельному типу. В старших гибридных потомствах, кроме сходных генотипов, выщепляются константные высокорослые формы, а также очень низкие трехгенные карлики. С. Ф. Лыфенко совместно с А. Мааткеримовым и Ю. Н. Пикой проанализировали отечественные и некоторые зарубежные полукарликовые и низкорослые сорта озимой пшеницы по их реакции на экзогенные ростовые вещества (гибберелловая кислота, 2, 4-Д и др.), что позволило в зависимости от функции генов карликовости разделить их на три группы: I—не реагирующие на ростовые вещества: Карлик 1, Одесская полукарликовая, Одесская 75, Лан, Обрий, Пересвет и др.; II— реагирующие на экзогенные ростовые вещества: Сава, Безостая 1, Одесская 51; III— сильно реагирующие, но их отзывчивость на ростовые вещества больше ингибируется светом (сорт Зирка).
Сорта последней группы сильно варьируют по признаку высоты растения. В зависимости от зоны выращивания, густоты стеблестоя и интенсивности инсоляции они могут быть полукарликами, низкорослыми или даже среднерослыми формами. Изучение генетической и физиолого-биохимической природы генов карликовости показало, что несмотря на их относительно большое количество (не менее 5), реакция всех генотипов на ростовые вещества сводится в основном к двум типам — отзывчивость и неотзывчивость, что обусловлено блокированием одного из двух процессов — синтез или утилизация рострегулирующих веществ.
Большинство генов карликовости обладает кумулятивным эффектом, что позволяет в зависимости от задач селекции «дозировать» признак. В настоящее время селекционер может располагать таким генетическим разнообразием по признаку высоты растения, которое позволяет по пшенице мягкой и твердой создавать практически любые сорта: карлики (30—50 см); полукарлики (55—80 см); низкорослые (85—100 см); среднерослые (105—140 см); высокорослые (более 140 см).
Укорочение стебля под влиянием генетических факторов сопровождается перестройкой его анатомической структуры. При этом наблюдаются большие изменения сосудистоволокнистых пучков и механических тканей, от которых в большой мере зависит не только устойчивость против полегания, но и продуктивность растений.
По наблюдениям К. Г. Тетерятченко, у сортов полукарликового типа сосудистоволокнистые пучки имеют более удачное строение и лучшие возможности усиления их основной физиологической роли — восходящего и нисходящего передвижения воды и продуктов фотосинтеза по стеблю. Большинство сортов полукарликового типа обладает более толстым стеблем, чем средне- и высокорослые сорта. Однако при сложных скрещиваниях с включением множества компонентов с разным диаметром и разной толщиной стенки соломины могут выщепляться очень тонкостебельные полукарликовые и карликовые формы, в том числе и такие, в стеблях которых практически отсутствует свободный канал из-за малого диаметра и относительно большой толщины стенки соломины. Эти формы интересны тем, что обладают неплохой устойчивостью против полегания и не повреждаются пилильщиком хлебным.
Некоторые селекционеры считают для степной засушливой зоны наиболее перспективными тонкостебельные высококустистые полукарликовые формы пшеницы. Однако большинство районированных в зоне сортов этого типа все же относится к толстостебельным формам. Наряду с положительными свойствами полукарликовые формы обладают отрицательными, которые приходится учитывать при решении селекционных задач. Одним из недостатков сортов полукарликового типа является короткое колеоптиле. Исследования С. Ф. Лыфенко и Н. И. Ериняка (1980), выполненные на разнообразных озимых и яровых полукарликовых сортах, показали очень четкую зависимость длины колеоптиле от высоты растений, коэффициент корреляции между этими показателями высокий (r=0,685±0,137). При этом высокая положительная корреляция оставалась постоянной и не зависела от типа и физиолого-биохимической природы гена карликовости. Зародышевые корешки по длине и массе почти не меняются в зависимости от генов, контролирующих высоту стебля (табл. 1).
Короткое колеоптиле является отрицательным свойством сорта. В случаях глубокой заделки семян и при недостатке в почве воды у форм с коротким колеоптиле наблюдается снижение полевой всхожести. Степень положительной корреляции между длиной колеоптиле и полевой всхожестью высокая (r=0,772±0,104).
Однако, несмотря на тесную связь этих признаков, в природе все же существуют полукарликовые формы с длинным колеоптиле. Так, яровой сорт мексиканской селекции Наинари 60 при высоте взрослого растения 60—65 см в период всходов имеет колеоптиле 9,5 см. Такая длина обычно бывает только у высоко- и среднерослых сортов (115— 140 см).
Короткостебельные формы существенно отличаются от обычных сортов по площади листовой поверхности, линейным размерам прикорневых и узловых листьев.
Исследования, проведенные на обычных и полукарликовых сортах озимой пшеницы, показали, что уже в фазе кущения гены карликовости каким-то образом влияют на размеры листовой поверхности, коэффициент корреляции между площадью листьев отдельных растений в фазе кущения и их высотой как генотипический признак, проявляющийся на заключительном этапе онтогенеза, довольно высокий (r=0,59±0,269). Наиболее интересным явлением оказалось то, что наследственные различия по высоте растения сказываются не только на формировании корреляционных связей в пределах отдельных растений, но и на общем фитоценозе посева. Причем коэффициент корреляции между высотой растений и листовым индексом (отношение площади листьев всего посева к занимаемой им площади поля) оказался очень высоким положительным (r=0,80+0,20). Следовательно, в процессе селекции короткостебельных форм пшеницы необходимо обращать внимание, в первую очередь, на генотипы с большой кустистостью и крупными листовыми пластинками, так как сорта интенсивного типа должны иметь хороший фотосинтетический потенциал. Несмотря на тесную положительную генотипическую корреляцию между высотой стебля и размерами листьев иногда удается гибридизацией получать короткостебельные формы с крупными листьями. Так, полукарликовый сорт Южная заря имеет листья такого же размера, как и среднерослый сорт Одесская 51. Однако большие размеры листьев в фазе кущения еще не свидетельствуют о большой урожайности сорта. Более того, сильное израстание растений в осенне-зимний и ранневесенний периоды зачастую приводит к уменьшению урожая. Коэффициент корреляции между листовым индексом в этот период и урожаем зерна достигает средней отрицательной величины (r=-0,37±0,23).
После выхода в трубку и колошения как у короткостебельных, так и обычных сортов в степной зоне часть листьев засыхает и теряет роль фотосинтезирующего органа. Однако интенсивность отмирания отдельных листьев и целых стеблей-подгонов неодинаковая. У низкорослых и полукарликовых форм, в отличие от обычных сортов, этот процесс протекает замедленно. В результате картина меняется на противоположную той, которая наблюдается в фазе кущения. У сортов полукарликового типа в проведенном опыте в фазе налива зерна листовой индекс составлял 3,49-4,96, в то время как у обычных сортов — 1,5-2. Коэффициент корреляции между листовым индексом этой фазы и урожаем зерна был положительным и довольно большим (r=0,5±0,11).
Таблица 1
Длинна колеоптиле и сила начально роста семян и форм озимой пшеницы мягкой с различным генов карликовости
| Группа генотипов по высоте растений | Количество генов карликовости | Высота взрослого растения, см | Длинна колеоптиле, см | Масса колеоптиле, г | Длинна корешков, см | Масса корешков, г |
| Среднерослые | Не более одного | 109,3 | 8,3 | 3,06 | 11,7 | 1,24 |
| Полукарликовые | Два | 60,5 | 6,1 | 2,04 | 11,0 | 1,30 |
| Карликовые | Три | 51,8 | 4,6 | 2,00 | 12,2 | 1,61 |
Приведенные данные показывают, что в процессе селекции короткостебельных сортов пшеницы необходимо обращать внимание на создание и отбор таких генотипов, которые способны более продолжительный период сохранять листья в активном жизнедеятельном состоянии. Листовой индекс в период налива зерна должен находиться в оптимальных границах 5-7. Полукарликовые сорта для юга Украины нужно, чтобы имели более низкие показатели облиственности, чем для центральной степной и лесостепной зон. Положение листьев (горизонтальный и прямостоячий, или торчащий типы) не зависит от функции генов карликовости. В процессе гибридного разнообразия с одинаковой частотой появляются высокорослые и короткостебельные формы с торчащим и горизонтальным листом. Исследования показали, что при уровне урожайности выше 35-40 ц/га предпочтительно иметь сорта с прямостоячей формой листа, который обычно соответствует этому уровню урожая, так как при стеблестое 450— 700 стеблей/м2 наблюдается взаимное угнетение стеблей и листьев. Средний урожай зерна сортов и селекционных форм в опытах с озимой пшеницей по черному пару был у форм с торчащим листом 55,68±1,69 ц/га (Р%=2,17), а с обычным — 52+0,79 ц/га (Р%=2,37). Таким образом, сорта с торчащим листом оказались урожайнее обычных на 3,68 ц/га. В связи с тем, что сорта короткостебельного типа предназначаются для интенсивной технологии, при которой формируется густой стеблестой, желательно, чтобы они, кроме короткого стебля, имели бы прямостоячий тип листа.
Проведенные экспериментальные исследования показали, что генотипической корреляции между высотой растения и размерами корневой системы не существует. Следовательно, имеется возможность селекционным путем создавать сорта короткостебельного типа с хорошо развитой корневой системой, не уступающие по этому признаку лучшим высокорослым сортам. Более удачное соотношение между размерами над- и подземной массы растений у сортов короткостебельного типа позволяет им в период налива зерна более экономно расходовать воду и успешнее противостоять почвенной и воздушной засухе, которая в этой фазе бывает особенно часто.
Признак короткостебельности связан со многими важными биологическими и хозяйственно ценными свойствами растений. Сорта высокоинтенсивного типа, в том числе низкорослые и полукарликовые, в большинстве случаев обладают меньшей морозостойкостью, чем степные сорта полуинтенсивного типа. Гены карликовости обладают плейотропным эффектом на многие биологические и морфологические признаки, с которыми связана устойчивость растений к действию отрицательных факторов периода зимовки. При этом эффекты генов карликовости неодинаково проявляются по отношению к морозостойкости и устойчивости против выпирания. Большинство изученных генов ослабляют морозостойкость, но повышают устойчивость против выпирания через заглубление узла кущения, которое осуществляется под влиянием укорочения эпикотиля — первого корневидного междоузлия. Характер генетической и физиолого-биохимической связи признака короткостебельности с морозостойкостью весьма сложный и остается до настоящего времени плохо изученным. В наибольшей степени с устойчивостью против низких температур связан тип онтогенетического развития растений.
В результате генетического анализа, выполненного отечественными и зарубежными исследователями с помощью моносомного метода, установлено, что гены, обуславливающие озимый образ жизни (яровизационный процесс), локализованы в хромосомах 2А; 2Д; 5А; 5В; 5Д и 7Д. В других исследованиях по тому же методу определено, что гены карликовости локализованы в хромосомах 1В, 7В и 5Д. Даже простое сопоставление результатов исследований показывает, что по меньшей мере одна хромосома (5Д) является одновременно ответственной за признак высоты растения и процесс яровизации. Имеются также сведения о том, что подобную роль играет и хромосома 2А. Следовательно, не только морозостойкость, но и тип развития растений могут иметь высокую степень генетического сцепления с признаком высоты растений. Однако в связи с полиморфизмом генов карликовости в процессе селекции имеется возможность создавать и отбирать из популяций такие короткостебельные генотипы, у которых этот признак имеет наименьшую отрицательную корреляцию с морозостойкостью.
Среди районированных и перспективных сортов полукарликового типа пока нет выдающихся по морозостойкости форм. Однако лучшие среди них приближаются по этому свойству к Одесской 51 и превосходит Безостую 1. Имеются среди них и недостаточно зимостойкие сорта (табл. 2).
Таблица 2
Морозостойкость полукарликовых сортов озимой пшеницы, % живых растений после промораживания при температуре 19°С (1981-1982 гг)
| Сорт | Морозостойкость | ||
| сорта | Одесской 51 | Безостой 1 | |
| Одесская полукарликовая | 72,7 | 89,4 | 48,2 |
| Одесская 75 | 71,0 | 89,4 | 48,2 |
| Зирка | 86,6 | 83,6 | 41,8 |
| Обрий | 47,6 | 81,5 | 46,0 |
| Южная заря | 75,5 | 83,6 | 41,8 |
| Лан | 62,3 | 89,4 | 48,2 |
| Бригантина | 72,5 | 81,5 | 46,0 |
| Донская полукарликовая | 70,5 | 81,5 | 46,0 |
Специальные исследования показывают, что путем направленных скрещиваний и отбора молено создать высокоморозостойкие сорта. Так, в ВСГИ получена полукарликовая селекционная линия Эритроспермум 2100/81, которая не уступает по морозостойкости Одесской 16— одному из наиболее зимостойких сортов озимой пшеницы. Признак высоты растения и функция отдельных генов карликовости связаны с накоплением запасных и функциональных белков в растении, что в последующем сказывается на технологических качествах зерна. Исследования, проведенные в ВСГИ А. Н. Хохловым, С. Ф. Лыфенко и Н. В. Тупицыным (1984), показали, что самый большой коэффициент хозяйственного использования у сортов полукарликового типа. У них соотношение зерна и соломы приближается 1:1. У средне- и высокорослых сортов урожай соломы больше, чем зерна. У настоящих карликов с высотой растения 35-50 см также урожай соломы больше, потому что зерно мелкое и плохо выполненное.
Запасные белки в зерне, от которых, в первую очередь, зависят его технологические свойства, как известно, формируются за счет двух взаимосвязанных процессов: 1— синтез аминокислот и прямая их транслокация в зерновку; 2 — реутилизация конституциональных белков всех вегетативных органов растения. В связи с тем, что у сортов полукарликового типа реутилизирующая масса (листья, стебли) меньше, чем у обычных сортов, соответственно меньше аминокислот поступает в зерновки за счет второго процесса. Кроме общих закономерностей, отмеченных в формировании технологических свойств зерна у короткостебельных сортов озимой пшеницы, существуют большие различия в плейотропном эффекте отдельных генов карликовости. В этом случае на первое место выступает специфичность разных генов карликовости. Так, гены карликовости, донором которых является сорт Норин 10, кроме уменьшения белка в зерне за счет увеличения коэффициента хозяйственного использования урожая, прямо не влияют на ухудшение качества зерна. В отличие от них ген карликовости краснодарского Карлика 1 обладает существенным отрицательным плейотропным эффектом на накопление белка и клейковины в зерне (табл. 3). Гены карликовости оказывают существенное влияние на количество клейковины в зерне, однако практически не меняют ее физических свойств. Многие сорта короткостебельного типа по показателям силы муки, устойчивости теста при замесе, объемному выходу хлеба и другим свойствам значительно превосходят лучшие сорта обычного типа. Отдельные полукарликовые сорта и по содержанию белка в зерне не уступают среднерослым сортам. Примером могут служить сорта Обрий, Ольвия, Прокофьевка и другие, которые имеют высоту растения 80-95 см, а по техническим свойствам зерна являются лучшими среди сортов озимой пшеницы. Они способны формировать урожай зерна сильной пшеницы даже в условиях орошения, где другие сорта редко дают продовольственное зерно. Короткостебельные формы пшеницы по основным биологическим свойствам не отличаются от обычных сортов этой культуры. В соответствии с этим все методы селекции, разработанные и проверенные на практике на обычных формах пшеницы, могут применяться и при создании сортов короткостебельного типа.
Вместе с тем короткостебельные формы, особенно их полукарликовая группа, обладают свойствами, которые обязательно следует учитывать практически на всех этапах селекции и в первичном семеноводстве. Характерным в селекционной работе с короткостебельными формами является то, что при гибридизации в качестве доноров генов карликовости зачастую приходится брать не лучшие широко распространенные в производстве сорта местной селекции, а коллекционные формы или сорта весьма отдаленного эколого-географического происхождения. Вызвано это двумя основными причинами. Во-первых, набор и генетическое разнообразие районированных и перспективных короткостебельных сортов пока остаются ограниченными. Во-вторых, трехгенные и в ряде случаев двухгенные карликовые формы сами по себе малопродуктивные и обладают комплексом других отрицательных свойств. При скрещивании с обычными сортами такие карлики могут давать одно- и двухгенные формы с оптимальной высотой растения 80—95 см. Так, от скрещивания трехгенного карлика Норин 10 в мире получено многочисленное количество короткостебельных сортов с одним и двумя генами карликовости. Такие же результаты в скрещиваниях могут дать карликовые сорта Олесен дварф, Зимбабве и ИР 301 из Индии, Су-вон (Южная Корея), Сончульзон (КНДР) и другие.
Таблица 3
Технологические качества зерна полукарликовых форм озимой пшеницы в сравнении с качеством обычных сортов при испытании по пару (1976 г.)
| Сорт, селекционная форма | Высота растения, см | Содер-жание белка в муке, % | Содержание сырой клейковины в муке, % | Удельная работа, дефор-мации теста, е.а. | Объем хлеба из 100 г муки, см3 | Пористость хлеба по 5-ти бальной шкале | Общая оценка хлеба по 5-ти бальной шкале |
| Обычные сорта | |||||||
| Одесская 51 | 118 | 14,2 | 30,2 | 263 | 890 | 3- | 3 |
| Безостая 1 | 118 | 13,7 | 27,8 | 258 | 940 | 4- | 4 |
| Эритроспермум 127 | 132 | 13,8 | 26,0 | 327 | 1100 | 4 | 4+ |
| Прибой | 120 | 13,1 | 25,6 | 180 | 1000 | 4- | 4+ |
| Кавказ | 123 | 14,4 | 29,5 | 199 | 850 | 3 | 3 |
| Одесская 66 | 115 | 14,6 | 26,2 | 250 | 930 | 3+ | 4- |
| Среднее по группе | 121 | 13,6 | 27,6 | 246 | 951 | 3+ | 4- |
| Полукарликовые сорта с мутантным геном карликовости от Карлика 1 | |||||||
| Эритроспермум 587/74 | 75 | 13,0 | 22,8 | 190 | 860 | 3 | 3 |
| Эритроспермум 892/74 | 75 | 13,1 | 22,2 | 268 | 940 | 2 | 3 |
| Эритроспермум 894/74 | 75 | 13,3 | 24,1 | 268 | 960 | 4- | 4 |
| Эритроспермум 245/74 | 75 | 13,0 | 23,5 | 273 | 950 | 3+ | 4 |
| Одесская 75 | 80 | 12,7 | 22,0 | 250 | 960 | 3+ | 3+ |
| Одесская полукарликовая | 75 | 13,9 | 25,6 | 223 | 880 | 3 | 3 |
| Лютесценс 893/74 | 75 | 12,4 | 22,7 | 211 | 1010 | 4 | 4- |
| Эритроспермум 260/74 | 80 | 13,0 | 23,2 | 258 | 970 | 4 | 4 |
| Эритроспермум 252/74 | 80 | 13,8 | 27,0 | 239 | 950 | 3 | 3+ |
| Среднее по группе | 76,7 | 13,13 | 23,8 | 262 | 942 | 3+ | 3+ |
| Полукарликовые формы с генами карликовости от Норин 10 | |||||||
| Обрий | 90 | 15,3 | 29,0 | 421 | 1010 | 5 | 5 |
| Эритроспермум 699/74 | 90 | 14,4 | 26,5 | 387 | 950 | 3+ | 4 |
| Эритроспермум 394/74 | 90 | 14,3 | 28,9 | 373 | 1050 | 4- | 4+ |
| Зирка | 85 | 14,8 | 30,0 | 293 | 1060 | 4+ | 5 |
| Среднее по группе | 88,8 | 14,7 | 28,6 | 368 | 1018 | 4+ | 5- |
Необходимость широкого использования форм иного географического происхождения ограничивает возможность применения принципов подбора пар для гибридизации по наименьшему количеству отрицательных свойств, а также по недостающим и взаимно дополняющим элементам продуктивности. Скрещивание экологически и географически отдаленных форм приводит к чрезвычайно широкому гибридному разнообразию и возникновению неожиданных форм в потомствах. Их появление позволяет иногда вывести селекцию культуры на качественно новый этап, например, создание сорта Безостая 1. Однако вероятность появления удачных рекомбинантов при скрещивании экологически и географически отдаленных форм очень незначительная. Чтобы увеличить ее, большинство селекционеров идут по пути увеличения количества гибридных комбинаций, иногда до 800-1500. Для сравнения отметим, что селекционные программы при создании обычных сортов выполняются на 50-120 комбинациях скрещивания.
Те же генетические закономерности, которые делают целесообразным увеличение количества комбинаций при скрещивании отдельных форм, создают необходимость увеличения количества потомств в Р2 и старших поколениях. При большом количестве гетерозиготных локусов желаемые рекомбинанты появляются с малой долей вероятности, а сам процесс расщепления продолжается очень долго. Некоторые задачи при гибридизации отдаленных форм удается решить только с применением сложных скрещиваний. Лучшие результаты обычно дают прерывающиеся замещающий и восстанавливающий беккроссы, а также ступенчатые скрещивания. При скрещивании высокорослых и короткостебельных форм пшеницы в Р2 и последующих гибридных поколениях наблюдается сложное расщепление по признаку высоты растений. В популяции появляются короткостебельные выщепенцы. Однако при густом посеве их можно потерять, так как они сильно угнетаются высокорослыми генотипами. Чтобы избежать этого, гибридные потомства следует высевать разреженным способом, обычно с междурядьями 45 см и с расстоянием в ряду 5-10 см.
Во время отбора родоначальных (элитных) растений следует обращать внимание на короткостебельные формы, если они даже значительно слабее высокорослых. Размноженные и посеянные в чистом виде короткостебельные формы очень часто бывают более продуктивными, чем высокорослые и среднерослые линии того же происхождения.
В связи с тем, что генетические свойства, определяющие морозостойкость и качество зерна, находятся в прямой зависимости от признака высоты растения, среди короткостебельных форм очень редко встречаются достаточно зимостойкие генотипы с хорошим технологическим качеством зерна. По этой причине приходится увеличивать вероятность их выделения, расширив масштабы селекционных посевов. Многие учреждения в нашей стране и за рубежом по программе создания короткостебельных сортов пшеницы имеют селекционный питомник пшеницы по 60-70 тыс., контрольный питомник — 3,5-4 тыс. линий.
Соответственно увеличивается количество лабораторных и лабораторно-полевых опытов но оценке селекционных форм на морозо-, зимостойкость и технологические свойства зерна.





Считаю фразу «…в частности при определении оптимальных доз рострегулирующих гербицидов…» не вполне корректной, поскольку гербициды призваны для уничтожения растений. Возможно, имелись ввиду ретарданты, или имелось ввиду подавляющее действие гербицидов, применяемых в посеве против сорняков, которое проявляется в ингибировании ростовых процессов
Татьяна, с Вами согласен, здесь что то не так. Информация взята с книги.
Здесь несколько вариантов, либо авторы имели ввиду «доз применяемых гербицидов», либо использовали слово «рострегулирующих» так как гербициды, которые применяются на пшенице, контролируют рост определенных сорняков.
Но думаю первый вариант более реальный, поэтому заменю на него.
Если у Вас есть какието другие варианты, пожалуйста пишите.